
TEIL I: ANATOMIE UND PHYSIOLOGIE
|
INHALT
DARSTELLUNG VON BAU UND FUNKTION DES NERVENGEWEBES
Das Nervensystem ist aufgebaut aus Nervenzellen, die meist vom umgebenden Gewebe durch spezielle "Stütz- oder Hüllzellen" (Gliazellen, Schwann- Zellen) abgegrenzt sind. Dabei bestehen zwischen den Nervenzellen Kontakte, sogenannte Synapsen.
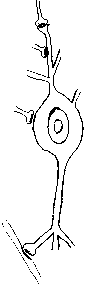 An den Enden dieser Zellfortsätze sind spezielle, knopfartige Verdickungen ausgebildet, die
mit anderen Nervenzellen in engem Kontakt stehen. Dabei kommen Kontaktstellen
von Ende zu Ende der Fortsätze, im Verlauf der Fortsätze oder vom Fortsatz
der einen zum Zelleib der anderen Nervenzelle vor. Unter starker Vergrößerung
zeigen die Kontaktstellen jeweils auf beiden Seiten besondere Strukturmerkmale:
es finden sich Auffaltungen und Verdickungen der Zellmembranen, im angrenzenden
Zytoplasma lagern kreisrunde Bläschen. Diese Kontaktstellen werden als
Synapsen bezeichnet und dienen dem Informationsaustausch zwischen den Nervenzellen.
An den Enden dieser Zellfortsätze sind spezielle, knopfartige Verdickungen ausgebildet, die
mit anderen Nervenzellen in engem Kontakt stehen. Dabei kommen Kontaktstellen
von Ende zu Ende der Fortsätze, im Verlauf der Fortsätze oder vom Fortsatz
der einen zum Zelleib der anderen Nervenzelle vor. Unter starker Vergrößerung
zeigen die Kontaktstellen jeweils auf beiden Seiten besondere Strukturmerkmale:
es finden sich Auffaltungen und Verdickungen der Zellmembranen, im angrenzenden
Zytoplasma lagern kreisrunde Bläschen. Diese Kontaktstellen werden als
Synapsen bezeichnet und dienen dem Informationsaustausch zwischen den Nervenzellen.

 Das PNS besteht im wesentlichen aus Nervenzellfortsätzen. Diese werden von der
Umgebung durch eine Umhüllung durch
die Schwann- Zellen getrennt. Dabei lassen sich die - meist dickeren -
Nervenzellfortsätze, die einzeln von mehreren Lagen zwiebelschalenartig
aufgerollten Schwann-Zellzytoplasmas umhüllt sind ("bemarkte Nerven",
links) von den - meist dünneren - Nervenzellfortsätzen unterscheiden,
die ohne "eigene" Schwann- Zelle zu mehreren zwischen Schwann-zellfortsätzen
verlaufen ("unbemarkte Nerven", rechts). Die Schwann-zellfortsätze
bilden dabei das sogenannte"Mark" (=Myelin), welches aufgrund
seines hohen Fettanteils (Phospholipide) bereits makroskopisch (mit bloßem Auge
sichtbar) den bemarkten
(=myelinisierten) Nerven ein weißlich-gelbliches Aussehen verleiht. Betrachtet
man einen bemarkten Nerven von der Längsseite, so lassen sich in regelmäßigen
Abständen Unterbrechungen der Markscheiden finden, die sogenannten Ranvierschen
Schnürringe. Sie entsprechen jeweils dem Übergang zweier Schwann- Zellen.
Eine große Anzahl von Fortsätzen verläuft gemeinsam und bildet im Bereich des
peripheren Nervensystems mit dem bloßen Auge sichtbare Faserzüge, die
peripheren Nerven.
Das PNS besteht im wesentlichen aus Nervenzellfortsätzen. Diese werden von der
Umgebung durch eine Umhüllung durch
die Schwann- Zellen getrennt. Dabei lassen sich die - meist dickeren -
Nervenzellfortsätze, die einzeln von mehreren Lagen zwiebelschalenartig
aufgerollten Schwann-Zellzytoplasmas umhüllt sind ("bemarkte Nerven",
links) von den - meist dünneren - Nervenzellfortsätzen unterscheiden,
die ohne "eigene" Schwann- Zelle zu mehreren zwischen Schwann-zellfortsätzen
verlaufen ("unbemarkte Nerven", rechts). Die Schwann-zellfortsätze
bilden dabei das sogenannte"Mark" (=Myelin), welches aufgrund
seines hohen Fettanteils (Phospholipide) bereits makroskopisch (mit bloßem Auge
sichtbar) den bemarkten
(=myelinisierten) Nerven ein weißlich-gelbliches Aussehen verleiht. Betrachtet
man einen bemarkten Nerven von der Längsseite, so lassen sich in regelmäßigen
Abständen Unterbrechungen der Markscheiden finden, die sogenannten Ranvierschen
Schnürringe. Sie entsprechen jeweils dem Übergang zweier Schwann- Zellen.
Eine große Anzahl von Fortsätzen verläuft gemeinsam und bildet im Bereich des
peripheren Nervensystems mit dem bloßen Auge sichtbare Faserzüge, die
peripheren Nerven.
Die Funktion des Nervensystem ist die Weitergabe und Verarbeitung von Informationen.
Es kommt sowohl zur
Differenzierung und Wertung zahlreicher Einflüsse wie auch zur Ausbreitung und
Generalisation einzelner Impulse. Das Grundprinzip der Informationsweitergabe
im Nervensystem sind elektrochemische Übertragungsvorgänge. Voraussetzung
sind die Fähigkeiten der Neurone, einen elektrischen Impuls zu empfangen
(elektrische Erregbarkeit), zu integrieren (Ruhepotential), gegebenenfalls
weiterzuleiten und zu übertragen (Aktionspotential, Erregungsleitung).
 Zwischen dem Inneren der Nervenzelle und der Umgebung besteht ein Spannungsunterschied,
das sogenannte
Ruhe(-membran-)potential. Im Normalzustand liegt die Spannung im Zellinneren
um 80 mV unter dem Zelläußeren, d.h. es besteht eine "Polarisation", welche
durch Konzentrationsunterschiede verschiedener geladener Teilchen ("Ionen")
zwischen Zellinnerem und Zellumgebung (Extrazellularraum) hervorgerufen wird.
Zwischen dem Inneren der Nervenzelle und der Umgebung besteht ein Spannungsunterschied,
das sogenannte
Ruhe(-membran-)potential. Im Normalzustand liegt die Spannung im Zellinneren
um 80 mV unter dem Zelläußeren, d.h. es besteht eine "Polarisation", welche
durch Konzentrationsunterschiede verschiedener geladener Teilchen ("Ionen")
zwischen Zellinnerem und Zellumgebung (Extrazellularraum) hervorgerufen wird.
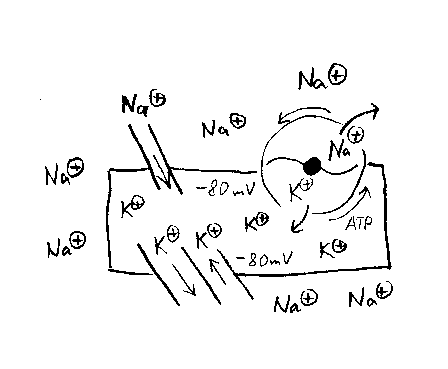
Die Zellmembran ist unterschiedlich durchlässig für verschiedenartige
Teilchen. Deshalb kommt es nicht zum Ausgleich der Konzentrationsunterschiede
dieser Teilchen zwischen Intra- und Extrazellularraum.
Bedeutsam für die Ausbildung des Ruhemembranpotentials ist
einerseits ein intrazellulärer Überschuss
negativ geladener, großer Anionen, überwiegend großmolekularer Eiweiße. Daneben
besteht ein starker intrazellulärer Überschuss positiv geladener Kalium-Ionen.
Im Ruhezustand können Kalium-Ionen die Membran relativ gut passieren, so das
entsprechend
dem Konzentrationsgefälle ein stetiger Kalium-Ausstrom existiert. Durch
den Verlust positiv geladener Teilchen bildet sich eine Ladungsdifferenz aus,
so das das Zellinnere gegenüber dem Extrazellularraum negativ geladen ist.
Diese Ladungsdifferenz wirkt allerdings einer weiteren Steigerung des Ausstroms
der positiv geladenen Kalium-Ionen entgegen, so das ab einer bestimmten
elektrischen Spannung die Kraft, die die Kalium-Ionen nach außen drängt
(Konzentrationsunterschied) einer gleichgroßen Kraft, die die Kalium-Ionen
in die Zelle zurückdrängt (Ladungsunterschied), gegenübersteht. Man spricht
von einem sogenannten Kalium-Gleichgewichtspotential, welches sich bei einem
Potential von etwa -80 mV ausbildet, dem sogenannten Ruhemembranpotential.
Gleichzeitig besteht eine erheblich geringe Permeabilität der Zellmembran
für Natrium-Ionen. In diesem Fall sind die treibenden
Kräfte - extrazellulär erhöhte Natriumkonzentration sowie extrazellulär erhöhte
Anzahl positiver Ladung - jedoch erheblich stärker, so das trotz der geringeren
Permeabilität ein merklicher kontinuierlicher Natriumeinstrom resultiert.
Beständiger passiver Kaliumausstrom und Natriumeinstrom würden zwar das
Membranpotential nicht verändern, es käme jedoch allmählich zu einem Ausgleich
der Teilchenkonzentrationen zwischen Intra- und Extrazellularraum. Dem
wirken energieabhängige sogenannte Ionenpumpen entgegen, welche aktiv jeweils
ein Kalium-Ion in die Zelle hinein, und dafür ein Natrium-Ion heraus pumpen.
Netto resultieren im Ruhezustand konstante Konzentrationsverhältnissen
der extra- und intrazellulären
Teilchen sowie ein konstantes Potential, das sogenannte Ruhemembranpotential.
 Neben den genannten Möglichkeiten, die Zellmembran zu passieren,
existieren in der Zellmembran hochspezialisierte
Öffnungen, durch die bei Bedarf Ionen strömen können und die spezifisch für
Kat- oder Anionen sind, bzw. oft nur eine ganz bestimmte Ionenart durchlassen.
Diese Öffnungen, die geöffnet und geschlossen werden können, bezeichnet man
als (Ionen-)Kanäle, es gibt u.a. sog. Natrium-, Kalzium- und Kaliumkanäle.
Neben den genannten Möglichkeiten, die Zellmembran zu passieren,
existieren in der Zellmembran hochspezialisierte
Öffnungen, durch die bei Bedarf Ionen strömen können und die spezifisch für
Kat- oder Anionen sind, bzw. oft nur eine ganz bestimmte Ionenart durchlassen.
Diese Öffnungen, die geöffnet und geschlossen werden können, bezeichnet man
als (Ionen-)Kanäle, es gibt u.a. sog. Natrium-, Kalzium- und Kaliumkanäle.
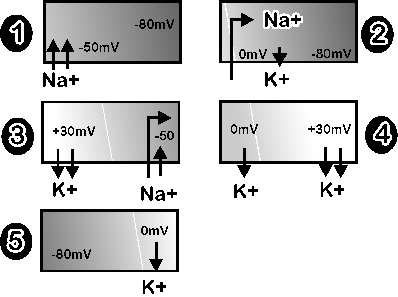
Bei einer elektrischen Erregung einer Nervenzelle kommt es zu einer Öffnung
der unterschiedlichen Ionenkanäle in einem genau festgelegten
zeitlichen Ablauf, beginnend mit Natriumkanälen, gefolgt von Kaliumkanälen.
Die in der Ausgangssituation bestehenden Konzentrationsunterschiede
der verschiedenen Ionenarten zwischen Zellinnerem und Extrazellularraum führen
dabei zu raschen Ionenverschiebungen entsprechend den Konzentrationsgefällen.
Nach Öffnung des Natriumkanals (Bild 1)gelangen im Überschuss
positiv geladene Natrium-Ionen ins
Zellinnere (Bild 2), woraus eine Änderung des Membranpotentials resultiert,
so das vorübergehend der Intrazellularraum gegenüber dem Extrazellularraum
um etwa +30 mV positiv geladen ist ("Depolarisation", Bild 3).
Bereits während (und als Folge) dieser Ladungsänderung schließt sich
zunächst der Natriumkanal. Durch anschließendes
Überwiegen der Kaliumpermeabilität dringen vermehrt positiv geladene
Kalium-Ionen nach außen (Bild 4, die "umgekehrte" Ladungsdifferenz
begünstigt dies zu diesem Zeitpunkt) und es kommt zum Wiederaufbau
des Ruhemembranpotentials ("Re-Polarisation", Bild 5), welches letztlich
wieder mit Hilfe der Ionenpumpen stabilisiert wird. Dieser gesamte, immer
gleichförmig ablaufende Vorgang wird als Aktionspotential bezeichnet.
Aktionspotentiale dauern bei den Nervenzellen 1 ms, an Muskelzellen 10 ms,
am Herzmuskel >200 ms.
Die zu Beginn des Aktionspotentials auftretende Öffnung der Natriumkanäle erfolgt durch
eine, meist von außen ausgelöste Änderung des Ruhemembranpotentials über einen
bestimmten Schwellenwert, der normalerweise bei -50 mV liegt ("Depolarisation").
Anschließend wird immer ein "vollständiges" Aktionspotential ausgelöst.
Es gilt das Alles-oder-Nichts-Prinzip, d.h.: wird der Schwellenwert erreicht,
findet ein typisches Aktionspotential statt, wird er nicht erreicht, passiert
nichts. Nach einem Aktionspotential kann der Natriumkanal vorübergehend
nicht durch Überschwelliege Depolarisation aktiviert werden, ein Aktionspotential
kommt nicht zustande. Die Zelle ist refraktär . Man unterscheidet
absolute Refraktärzeit (bei Nervenzellen 1-2 ms) und relative Refraktärzeit, in
der eine verstärkte Depolarisation doch noch ein Aktionspotential auslösen kann.
Durch verschiedene Maßnahmen wie Medikamente, Konzentrationsänderungen
der Ionen oder eine außerordentlich
langsame Annäherung an diese Schwelle lässt sich der Schwellenwert
allerdings verschieben, durch Sauerstoffmangel oder mit Hilfe bestimmter
Medikamente (Muskelrelaxantien) kann die Aktivierung des Netriumkanals
sogar vollständig inaktiviert und ein Aktionspotential somit verhindert werden.
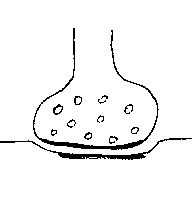 Eine elektrische Übertragung über den synaptischen Spalt ist nicht möglich.
Das an der präsynaptischen Membran eintreffende Aktionspotential
führt dazu, das dort in sogenannten synaptischen Bläschen gespeicherte
Übertragungsstoffe (=Transmitter) in den synaptischen Spaltraum
freigesetzt werden. Im Normalfall finden sich in der postsynaptischen Membran
sogenannte Rezeptorstrukturen, die genau für diesen Transmitter zugeschnitten
sind und beim Eintreffen des Transmitters "ihrer" (postsynaptischen)
Zelle signalisieren, das ein Erregungsvorgang stattgefunden hat. Bei der
postsynaptischen Zelle kann es sich nun wiederum um eine Nervenzelle bzw. den
Dendriten einer Nervenzelle, genauso gut jedoch auch um eine Muskelzelle oder
z.B. eine Schweißdrüsenzelle handeln. Je nachdem wird eine bestimmte Funktion
ausgelöst: die Nervenzelle könnte die Erregung wiederum weiterleiten, die
Muskelzelle sich zusammenziehen oder die Schweißdrüsenzelle Schweiß produzieren.
Eine elektrische Übertragung über den synaptischen Spalt ist nicht möglich.
Das an der präsynaptischen Membran eintreffende Aktionspotential
führt dazu, das dort in sogenannten synaptischen Bläschen gespeicherte
Übertragungsstoffe (=Transmitter) in den synaptischen Spaltraum
freigesetzt werden. Im Normalfall finden sich in der postsynaptischen Membran
sogenannte Rezeptorstrukturen, die genau für diesen Transmitter zugeschnitten
sind und beim Eintreffen des Transmitters "ihrer" (postsynaptischen)
Zelle signalisieren, das ein Erregungsvorgang stattgefunden hat. Bei der
postsynaptischen Zelle kann es sich nun wiederum um eine Nervenzelle bzw. den
Dendriten einer Nervenzelle, genauso gut jedoch auch um eine Muskelzelle oder
z.B. eine Schweißdrüsenzelle handeln. Je nachdem wird eine bestimmte Funktion
ausgelöst: die Nervenzelle könnte die Erregung wiederum weiterleiten, die
Muskelzelle sich zusammenziehen oder die Schweißdrüsenzelle Schweiß produzieren.
Eine Besonderheit ergibt sich bei der synaptischen Übertragung zwischen
Nervenzellen: eine synaptische Übertragung löst bei der postsynaptischen
Zelle NICHT automatisch nach dem Alles-oder-Nichts-Prinzip erneut
ein Aktionspotential aus. Stattdessen führt eine einzelne Übertragung entweder
zu einer Teil-Depolarisation der Zellmembran, so das ein neues Aktionspotential
erst nach der nächsten oder einer späteren Erregungsübertragung,
eben nach Erreichen der Depolarisationsschwelle zustande kommt. Daneben
gibt es auch Synapsen, die das Gegenteil bewirken: eine Erregungsübertragung
löst hier eine verstärkte Polarisation der postsynaptischen
Zelle aus, so daß ein Zustandekommen eines Aktionspotentials dieser Zelle sogar
noch erschwert wird. Eine solche Synapse wird als hemmend (=inhibitorisch),
die zuvor beschriebene Art als erregend (=excitatorisch) bezeichnet.
An den Dendriten und den Zelleibern der meisten Nervenzellen findet
sich ein Gemisch aus inhibitorischen und excitatorischen Synapsen. Der
jeweilige Erregungszustand solcher Nervenzellen stellt demnach eine Integration
der aus unterschiedlichen Richtungen eingetroffenen Informationen dar.